
表的内容
- β-氧化是分解法的过程脂肪酸分子在原核生物的细胞质中被分解,在真核生物的线粒体中被分解,生成乙酰辅酶a。
- 乙酰CoA进入三羧酸循环虽然nadh和fadh2(即共酶)用于电子传输链。
- 它被称为“氧化”,因为脂肪酸中的碳经过氧化生成羰基。
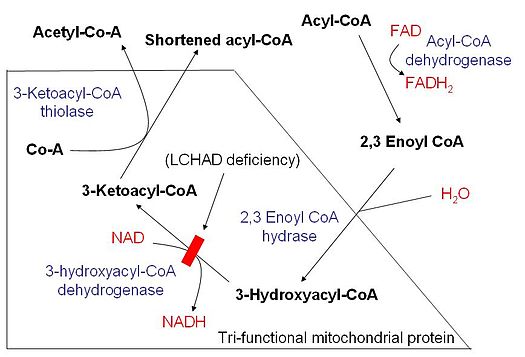
图:示意图显示线粒体脂肪酸β-氧化和长链3-羟基乙基 - 辅酶的脱氢酶缺乏,LCHAD缺乏症。资料来源:维基百科。
位置的机会
β-氧化在真核生物的线粒体中发生,而在原核生物中的细胞溶胶中。
- 基板:游离脂肪酸;H2O.
- 产品:一个乙酰辅酶a,一个NADH,一个FADH2用于从脂肪酸链中除去两碳基团。
氧化途径
- 在线粒体中,脂肪酸经历一系列氧化和水合反应,这导致从脂肪酸链中除去两碳基团(以乙酰COA形式)以及一个NADH和一个形成FADH.2,进入电子传输链以形成五个ATP。
- 形成的乙酰CoA将进入柠檬酸循环,然后进入电子传输链,导致形成另外12个ATP。循环继续,每圈除去另外的两碳基团,直到原始的长链脂肪酸还原为乙酰COA或丙酰基COA。
- 丙酰基CoA可以通过三种酶活性转化为琥珀酰CoA,其需要生物素和维生素B12作为辅因子,然后琥珀酰CoA可以进入柠檬酸循环。
A.活化脂肪酸
- 在细胞的细胞溶溶胶中,通过ATP和辅酶A激活长链脂肪酸,形成脂肪酰基CoA。短链脂肪酸在线粒体中激活。
- ATP转化为AMP和焦磷酸盐(PPI),其通过焦磷酸酶裂解至两种无机磷酸盐(2 PI)。因为两种高能磷酸键粘合,所以两种ATP分子用于脂肪酸活化。
B.脂酰辅酶a从细胞质到线粒体的转运
- 来自胞质的脂肪酰基辅酶a与线粒体外膜中的肉碱反应,形成脂肪酰基肉碱。这种酶是肉碱酰基转移酶I (CAT I),也叫肉碱棕榈酰转移酶I (CPT I)。脂肪酰基肉碱通过内膜,在那里重新形成脂肪酰基辅酶a,进入基质。第二种酶是肉碱酰基转移酶II (CAT II)。
- 催化来自辅酶A至肉碱的丙氨酸酰基转移酶I通过丙二酰基抑制脂肪酸合成中中间体抑制。因此,当在胞质溶溶胶中合成脂肪酸时,丙二酰库抑制其转运进入线粒体,因此防止了徒劳的循环(合成,然后立即降解)。
- 在线粒体内部,脂肪酰基辅酶a经历-氧化。
C.偶链脂肪酸的β氧化
β-氧化(其中所有反应涉及脂肪酰基-CoA的β-碳)是由四个顺序步骤组成的螺旋,其前三个类似于琥珀酸盐和草乙酸酯之间的TCA循环中的螺旋。重复这些步骤,直到偶数链脂肪酰基-CoA的所有碳转化为乙酰辅酶。
- FAD在第一步从脂肪酸辅酶a中接受氢。α-和β-碳之间产生双键,形成烯酰辅酶A。的FADH2与电子传递链相互作用,产生ATP。
- 酶:酰基CoA脱氢酶(这种酶的多种变体)
- H2o在双键上增加,形成β-羟基乙基-COA。
- 酶:Enoyl-CoA水合酶
- 通过NAD +氧化β-羟基酰基-COA至β-酮酰基-COA。产生的NADH与电子传输链相互作用,产生ATP。
- 酶:L-3-hydroxyacyl-CoA脱氢酶(对β-羟基酰基辅酶a的l-异构体特异)。
- β-酮酰基-CoA的α和β碳之间的粘合由需要辅酶A.乙酰-CoA由原始脂肪酰基-Coa的羧基末端的两种碳产生,以及剩余的碳形成脂肪酰基 - COA,这是比原来短的两条碳。
- 酶:β-酮硫酶
- 缩短的脂酰辅酶a重复这四个步骤。重复进行,直到原来的脂酰辅酶a的所有碳都转化为乙酰辅酶a。
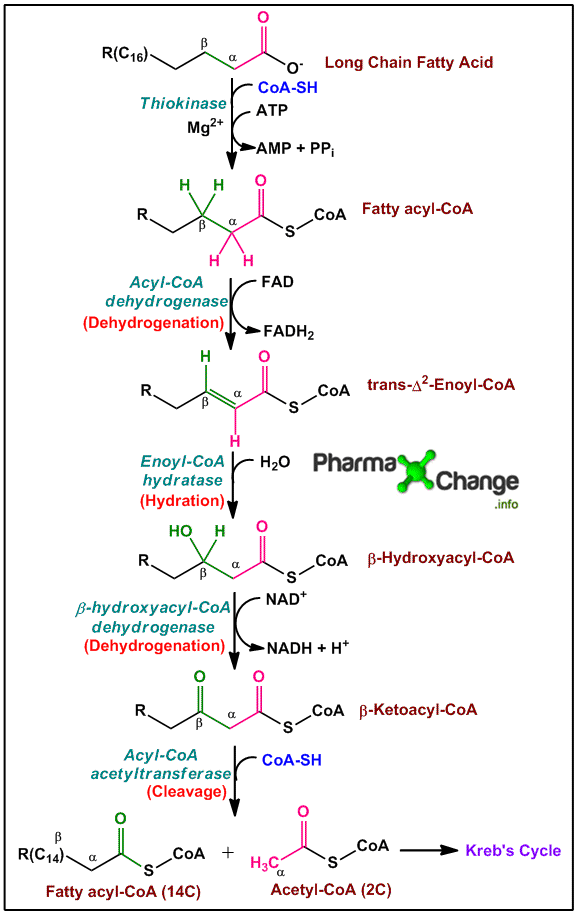
图:甚至碳链长度β氧化饱和脂肪酸。资料来源:Pharma X改变
均匀链脂肪酸的能量产率
- 能量是由β氧化产物产生的。
- 16碳棕榈酰辅酶a经过7个重复。
- 在最后一个重复中,一个4-碳脂肪酸酰辅酶a(丁基辅酶a)被裂解为两个乙酰辅酶a。
1.当一个Palmitoyl-CoA被氧化时,七个FADH2,形成七个NADH和八个乙酰COA。
- 七个fadh.2每次产生大约1.5个ATP,总共产生大约10.5个ATP。
- 7个NADH各产生约2.5个ATP,共产生约17.5个ATP。
- 这8个乙酰辅酶a可以进入TCA循环,每个产生约10个ATP,总共约80个ATP。
- 从棕榈酰-COA氧化到CO2和H2O,产生约108ATP。
2.从血液中进入血液中的棕榈酸的净ATP是约106,因为在可以氧化之前,棕榈酸酯必须经过活化(需要相当于2 ATP的过程)(108 ATP - 2 ATP = 106 ATP)。
3.其他脂肪酸的氧化将产生不同量的ATP。
D.奇数链和不饱和脂肪酸的氧化
- Odd-chain脂肪酸产生乙酰辅酶a和丙酰辅酶a。
- 这些脂肪酸重复β-氧化螺旋的四个步骤,产生乙酰辅酶a,直到最后一次裂解,剩下的三个碳以丙酰辅酶a的形式释放出来。
- 丙酰辅酶a,而不是乙酰辅酶a,可以转化为葡萄糖。
- 不饱和脂肪酸包括大约一半的人脂质中的脂肪酸残基,除了催化β-氧化螺旋的重复步骤之外还需要酶。
- 反应途径根据双键是否处于偶数或奇数碳位置而不同。
- β -氧化发生,直到不饱和脂肪酸的双键靠近脂肪酸酰基链的羧基端。
(1)如果双键起源于奇数碳数(例如3,5,7等),则异构酶将最终的顺式δ3转化为反式δ2脂肪酸。
(2)如果双键起源于均匀碳数(例如4,6,8等),则最终的反式δ2,顺式-δ4脂肪酸将通过2,4-二烯酰基降低CoA还原酶需要NADPH并产生反式δ3-酰基-COA和NADP1。异构酶将转换反式δ3脂肪酰基-COA转换为反式δ2脂肪酰基-CoA以允许β-氧化继续。
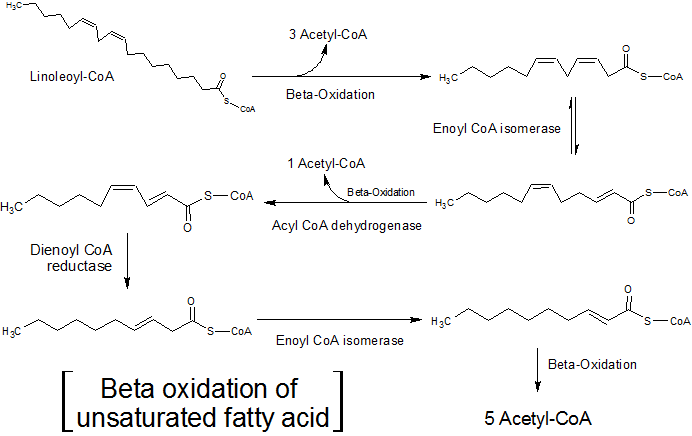
资料来源:维基百科
不饱和脂肪酸的ATP产率
(1)如果双键起源于奇数碳位置,则与相同碳长度的完全饱和脂肪酸相比,由于生产的FADH2较少,因此在奇数碳位置下的每个不饱和度将存在1.5 ATP对于每个不饱和度。
(2)如果双键起源于偶数碳位置,那么与等长完全饱和脂肪酸相比,由于在2,4-二烯基辅酶a还原酶催化的步骤中使用了NADPH,会少产生一个NADH当量(或2.5 ATP)。
β氧化的总体反应
一个氧化循环的总反应是:
CN-酰基CoA + FAD + NAD++ H2O + CoA→CN-2-酰基CoA + FADH2+ nadh + h++乙酰辅酶a
重要的酶
- 酰基CoA脱氢酶:在脂肪酸链的α和β碳原子之间形成双键。产生一个FADH2。
- Enoyl辅酶a水合酶:将水分子加入脂肪酸链中,从而打破α和β碳原子之间的双键。
- 3-羟基 - 酰基COA脱氢酶:再次将脂肪酸链脱氢,从而在β碳和氧分子之间形成一个双键。产生一个NADPH。
- 酰基COA酰基转移酶:在脂肪酸链的末端切割乙酰CoA,加入β碳。
调节脂肪酸的β-氧化
通过控制氧化磷酸化的机制来调节β-氧化(即,通过对ATP的需求)的调节。
- 催化剂:肾上腺素通过激活营养依赖性蛋白激酶刺激β-氧化,这导致磷酸化,从而激活HSL。当活化时,HSL从脂肪组织中释放脂肪酸和甘油以进行β-氧化。
- 抑制剂:胰岛素通过去磷酸化HSL抑制β-氧化,从而抑制来自脂肪组织的脂肪酸的释放。
意义
脂肪酸是人体中的主要能量来源,主要通过β-氧化氧化。
参考
- 史密斯,c.m.,马克斯,a.d.,利伯曼,m.a.,马克斯,d.b., &马克斯,d.b.(2005)。马克斯的基础医学生物化学:临床方法。费城:Lippincott Williams & Wilkins。
- 林宁格,A. L.,纳尔逊,D. L.,考克斯,M. M.(2000)。生物化学的Lehninger原理。纽约:Worth Publishers。
- John W.Pelley,爱德华F. Goljan(2011)。生物化学。第三版。费城:美国。
如何引用你的参考。我想引用您的信息。
感谢您。
你可以引用Aryal S. 2018.脂肪酸的β-氧化。微生物笔记。从://www.kirikcitarim.com/beta-oxidation-of-fatty-acid/