
目录
经典途径是一种事件链,其中补体组分在特定序列中反应作为级联导致细胞裂解。它由与抗原结合的抗体激活(抗原 - 抗体复合物),但不是由天然或游离抗体。这些配合物可以是可溶的,或者可以在抗体与抗原决定因素或表位结合时形成,或者在位于病毒,真菌,寄生或细菌细胞膜上。可溶性抗体 - 抗原复合物通常被称为免疫复合物,只有复合物形成IgM或IgG的某些子类抗体能够激活经典补体途径。
古典途径的激活剂
古典途径的激活剂包括以下内容:
- 免疫球蛋白IgM和IgG。IgG子类关于它们在激活补体方面的效率方面变化;IgG3是最有效的,其次是IgG1和IgG2。IgG4不要激活经典路径。
- 葡萄球菌蛋白A,
- C-反应蛋白
- 脱氧核糖核酸
古典途径激活的步骤
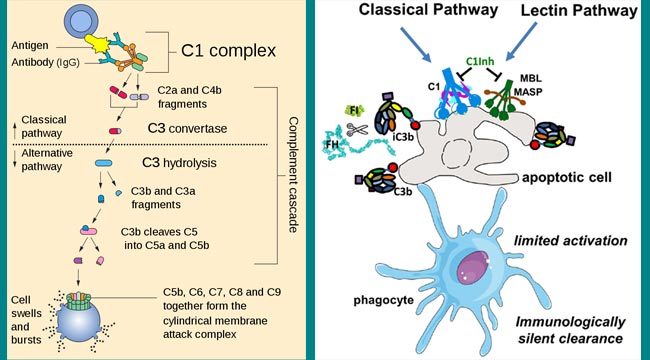
补体激活的经典途径通常以可溶性抗原 - 抗体复合物(免疫复合物)的形成或与抗体与抗原的结合在合适的靶标,例如细菌细胞上。以下是古典途径激活的顺序步骤:
- C1的激活:活化的初始阶段涉及补体组分C1,C2,C3和C4,其存在于血浆中作为酶的血浆。抗原抗体复合物的形成诱导抗体分子的非抗原结合(Fc)部分的构象变化。这种构象变化暴露了C1组分的结合位点。这导致C4,C2和C3的顺序激活。
在血清中,C1作为大分子复合物,其由一个C1Q和两个分子中的两种分子组成,每种丝氨酸蛋白酶,C1R和C1s,在Ca中保持在一起++稳定的复合物(C1QR2S2)。C1Q分子本身由18个多肽链组成,其与形成六种胶原状三螺旋臂的缔合物,其尖端结合抗原结合抗体分子的CH2结构域。每个C1大分子复合物必须由其C1Q球形头与至少两个Fc位点结合,以进行稳定的C1-抗体相互作用
- 在钙离子存在下C1Q结合导致C1R和C1s的活化。C1Q与CH的绑定2抗原复合抗体分子的Fc区的结构域在将其转化为活性丝氨酸蛋白酶的C1R分子中的一个构象变化。然后该C1R分子切割并激活其合作伙伴C1R分子。然后,两个C1R蛋白酶切割并激活两个C1S分子。活化的C1s是酯酶,其将C4分成两个片段:小可溶性片段(C4a)和较大的片段(C4b)。C4a具有过敏反应活性,C4b与细胞膜与C1结合。C4b在mg存在下++将C2分成C2A和C2B。较小的片段(C2b)扩散,而较大的片段(C2a)保持在C4b上。得到的C4b2a复合物具有酶活性并且被称为C3转换酶,将C3转换为活动形式。
- C3转化酶激活数千个C3分子,并将这些分子分成C3A和C3B。单个C3转化酶分子可以产生超过200个C3b分子,在该序列的该步骤中产生巨大的扩增。活化C3b以及C4B的生物重要性是它们能够结合C3B / C4B受体(目前指定为CR1受体)几乎所有宿主细胞,最符合吞噬细胞。吞噬细胞对C3B(或IC3B)/ C4B涂覆颗粒的增加的亲和力是已知为的免疫粘附。后者负责吞噬作用的显着增强,这是身体的主要防御机制之一。
- 一些C3b与C4B2A结合,形成称为分子复合C4b2a3bC5转换酶。C5转化酶将C5分成C5a和C5b。C5a扩散,而C5b附着到C6并引发C5B-9复合物的形成,否则称为膜攻击复合体(Mac)。
注意点:
当C1S从其其中一个链的氨基末端水解小片段(C4a)时,激活C4。C4b片段在C1附近共价成共价至靶膜表面,然后结合C 2。当C4b上暴露在C4裂解时不稳定的内部硫酯时,发生与膜结合的C4b发生结合,与蛋白质的羟基或氨基的羟基或氨基反应,或在细胞膜上反应。这反应必须快速发生,否则硫酯C4b进一步水解并可不再与细胞表面进行共价键。
只有与抗原结合的抗体,而不是自由循环抗体,可引发经典途径激活。原因是每个C1Q分子必须与至少两个待活化的IG重链结合,并且每个Ig Fc区仅具有单个C1Qbinding位点。因此,C1必须可以访问两个或更多个FC区以启动经典途径激活。
良好的信息,谢谢
太感谢了
我总是喜欢阅读你的文章,它是如此丰富和教育
太感谢了
你的写作总是丰富和教育。
太感谢了