
目录
真核转录
- 转录是将DNA链中的信息复制到新分子中的过程RNA..
- 它是基因表达的第一步,其中通过酶RNA聚合酶将特定的DNA片段复制到RNA(尤其是mRNA)中。
- 它会产生一种互补的、反平行的RNA链,称为初级转录本。
在真核生物中转录
在真核生物中转录发生的方式与原核生物类似,涉及的基本步骤。然而,它们之间的一些主要区别包括:
- 发起更复杂。
- 终止不涉及茎环结构。
- 转录由三种酶(RNA聚合酶I,II和III)进行。
- 转录的调节比原核生物更广泛。
参与真核生物转录的酶
不像原核生物,所有的RNA都是由一个RNA聚合酶合成的,真核细胞的细胞核有三个RNA聚合酶负责转录不同类型的RNA。
- RNA聚合酶I(RNA POL I)位于核仁中,并转录28s,18s和5.8s rRNA基因。
- RNA聚合酶II (RNA Pol II)位于核质中转录蛋白编码基因,产生pre-mRNA,同时编码参与rRNA加工的小核仁rna (small nucleolar RNAs, snoRNAs)和参与mRNA加工的小核rna (small nuclear RNAs, snRNA)的基因,U6 snRNA除外。
- RNA聚合酶III(RNA POL III)也位于核状物中。它转录TRNA,5S rRNA,U6 SnRNA的基因,以及与在内质网膜上易位的信号识别粒子(SRP)相关的7S RNA。
- 这三种真核RNA聚合酶中的每一种都包含12个或更多的亚基所以它们是大型的复杂酶。
- 编码每个真核酶的某些亚基的基因与编码核心酶的亚基的基因的DNA序列相似大肠杆菌RNA聚合酶。
- 然而,每个真核RNA聚合酶的其他4 - 7个亚基是独特的,它们与细菌RNA聚合酶亚基或其他真核RNA聚合酶的亚基没有相似之处。
真核生物转录的特征
- 真核生物的转录发生在细胞核内,mRNA从细胞核进入细胞质进行翻译。
- RNA聚合酶的RNA合成起始是由转录起始位点5 '侧的启动子位点引导的。
- RNA聚合酶转录DNA模板的一条链,反义( - )链。
- RNA合成不需要引物。
- RNA合成发生在5 '→3 '方向,RNA聚合酶催化生长中的RNA链的3- oh在进入的5-三磷酸核糖核苷上的-磷原子上进行亲核攻击。
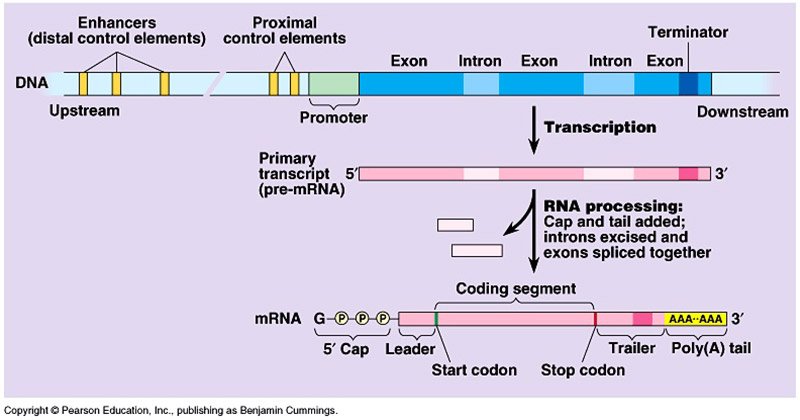
- 真核生物中的mRNA是从初级RNA转录物处理的,该方法称为成熟。
真核生物转录过程
这些真核RNA聚合酶合成RNA的基本机制可以分为以下几个阶段:
启动阶段
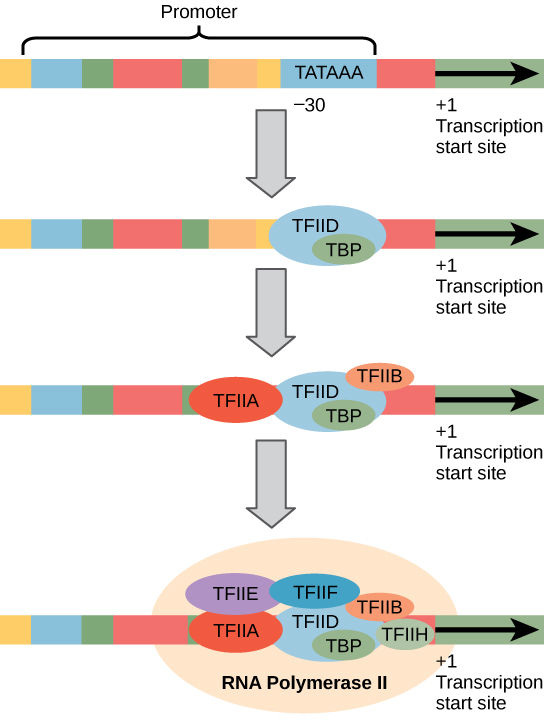
- 在起始阶段,RNA聚合酶识别DNA上一个特定的位置,位于将要转录的基因上游,称为a启动子站点然后在本地展开DNA。
- Most promoter sites for RNA polymerase II include a highly conserved sequence located about 25–35 bp upstream (i.e. to the 5 side) of the start site which has the consensus TATA(A/T)A(A/T) and is called the TATA box.
- 由于起始位置被表示为位置+1,TATA盒位置被称为大约位于位置-25。
- TATA框序列类似于代理中的-10序列(TATAAT),除了它位于上游进一步。
- 两种元素具有基本相同的功能,即由RNA聚合酶识别,以便在正确的位置定位酶以引发转录。
- 塔塔盒周围的顺序也很重要,因为它影响起始效率。转录也由位于TATA盒5 '的上游控制元件调控。
- 一些真核蛋白质编码基因缺乏塔塔盒,并替代是引发元件,以转录引发位点为中心。
- 为了启动转录,RNA聚合酶II需要几个其它蛋白质或蛋白质复合物的协助,这些蛋白质或蛋白质复合物被称为一般(或基础)转录因子,它们必须在启动子上组装成一个复合物,以便RNA聚合酶结合并开始转录。
- 这些都有TFII (RNA聚合酶II的转录因子)的通用名称。
- 起始阶段的第一个事件是转录因子IID (TFIID)蛋白复合物通过一个称为TBP (TATA box binding protein)的亚基与TATA box结合。
- 一旦TFIID复合物绑定,TFIIA结合并稳定TFIID-TATA盒相互作用。接下来,TFIIB绑定到TFIID。
- 然而,TFIIB也可以结合RNA聚合酶II,因此作为桥接蛋白。因此,
- 已经与TFIIF结合的RNA聚合酶II现在结合。
- 接下来是TFIIE和h的结合。这个最终的蛋白质复合物包含至少40个多肽,被称为转录起始复合物。
- 那些有启动元件而不是TATA盒的蛋白质编码基因似乎需要另一个与启动元件结合的蛋白质。
- 然后其他转录因子结合形成转录起始复合物,其方式与上面描述的具有TATA盒启动子的基因类似。
伸长阶段
TFIIH有两个功能:
- 它是一种螺旋酶,这意味着它可以使用ATP来放松DNA螺旋,允许转录开始。
- 此外,它磷酸化RNA聚合酶II,使该酶改变其构象,并与起始复合物中的其他蛋白质分离。
- 关键的磷酸化发生在RNA聚合酶II分子的c端长尾,称为c端结构域(CTD)。
- 有趣的是,只有具有非磷酸化CTD的RNA聚合酶II只能引发转录,而是只有RNA聚合酶II,具有磷酸化的CTD可以伸长RNA。
- RNA聚合酶II现在开始沿着DNA模板移动,合成RNA,也就是说,这个过程进入延伸阶段。
- RNA合成发生在5 '→3 '方向,RNA聚合酶催化生长中的RNA链的3- oh在进入的5-三磷酸核糖核苷上的-磷原子上进行亲核攻击。
- 由RNA聚合酶II由蛋白质编码基因制成的RNA分子称为初级转录物。
终止阶段
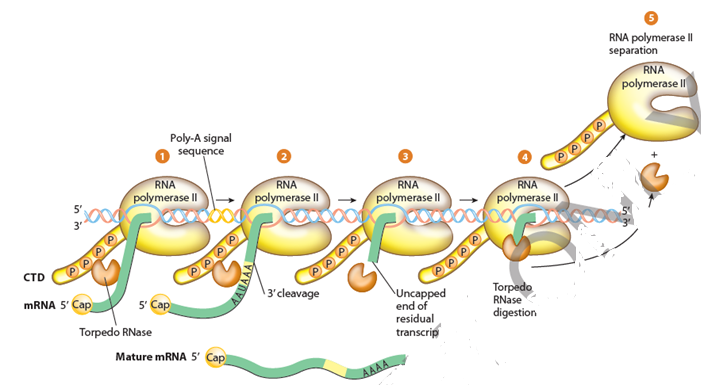
- RNA链的延伸一直持续到终止发生。
- 与原核生物中的RNA聚合酶不同,RNA聚合酶II不会在特定位点终止转录,而是可以在基因下游不同距离终止转录。
- 通过RNA聚合物II转录的RNA基因缺乏任何特定信号或序列,即直接RNA聚合酶II终止于特定位置。
- RNA聚合酶II可以继续从几千BP到数千名BP的任何地方转录RNA,超过基因的实际结束。
- 转录物在RNA聚合酶II完成转录之前在内部部位切割。这释放了转录物的上游部分,其在进一步处理之前将用作初始RNA(在蛋白质编码基因的情况下PRE-mRNA。)
- 该切割位点被认为是基因的“结束”。转录物的其余部分通过5'-外切核酸酶(在人体内称为XRN2)消化,而其仍然被RNA聚合酶II转录。
- 当通过消解所有悬垂的RNA时,5'-外含油酶“捕获到RNA聚合酶II时,它有助于将聚合酶与其DNA模板链脱离,最终终止该转录。
RNA加工
原真核生物mRNA转录较长,且定位于细胞核内,也称为异质核RNA (hnRNA)或pre- mRNA。
它经历了不同的加工步骤,变成成熟的RNA:
分裂
- 较大的RNA前体被切割成形以形成较小的RNA。
- 初级转录本被核糖核酸酶- p(一种RNA酶)切割,形成5-7 tRNA前体。
限制和尾矿
- 最初在5 '端加入一个帽(由7-甲基鸟苷或7 mG组成)和poly a的尾巴在3 '端。
- 该帽是一种经化学修饰的鸟苷三磷酸(GTP)分子。
拼接
- 真核生物初级mrna由两种类型的片段组成;非编码内含子和编码外显子。
- 内含子通过一种叫做RNA剪接的过程被移除,ATP被用来剪切RNA,释放内含子并连接两个相邻的外显子来产生成熟的mRNA。
核苷酸的修改
- 它们是TRNA - 甲基化(例如甲基胞嘧啶,甲基胍),脱氨基(例如来自腺嘌呤的Inosine),Dihydratacil,假尿嘧啶等中的最常见的。
转录后处理需要将初级转录本转化为功能性rna。
意义
- DNA的转录是调节基因表达的方法。
- 它发生在蛋白质翻译的准备过程中,也是蛋白质翻译所必需的。
参考文献
- 大卫·海姆斯和奈杰尔·胡珀(2005)。生物化学。第三版。泰勒和弗朗西斯集团:纽约。
- Bailey, W. R., Scott, E. G., Finegold, S. M., & Baron, E. J.(1986)。贝利和斯科特的诊断微生物学。圣路易斯:处于。
- Madigan, M. T., Martinko, J. M., Bender, K. S., Buckley, D. H., & Stahl, D.(2015)。《布罗克微生物生物学》(第十四版)。波士顿:皮尔森。
- http://www.biologydiscussion.com/rna/transcription/transcription-in-prokaryotes-and-eukaryotes-with-diagram/15546
- https://courses.lumenlearning.com/boundless-biology/chapter/eukaryotic-transcription/