
目录
- 翻译是指在蛋白质合成过程中,将信使RNA (mRNA)分子的序列翻译成氨基酸序列。
- 它是DNA转录为RNA后,细胞质或内质网中的核糖体合成蛋白质的过程。
核糖体
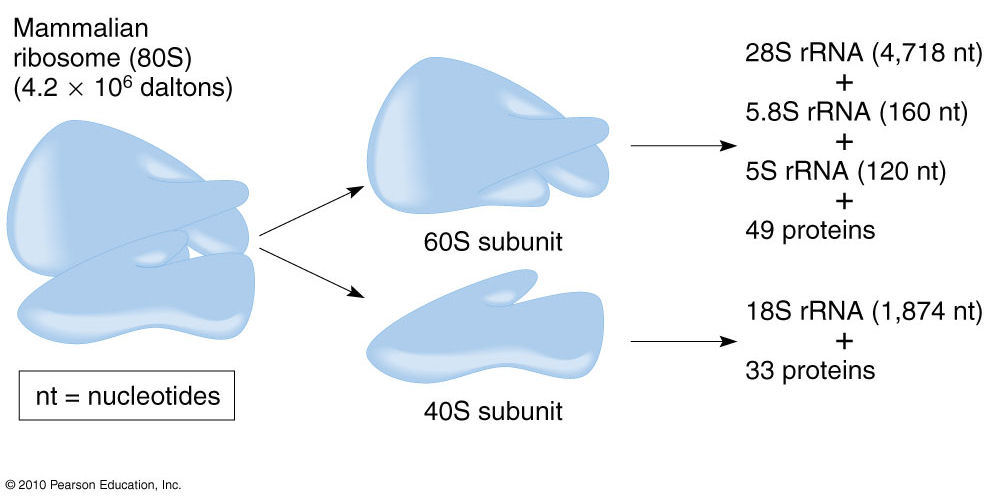
- 核糖体通常作为独立的亚基存在,由蛋白质和rRNA组成。
- 真核核糖体较大(80秒)和比原核性核糖体(70s)更复杂。
- 当亚基结合到mRNA的5 '端附近时,它们聚集在一起形成核糖体。
- 在与mRNA结合时,核糖体从5'至3'方向读取核苷酸序列,将来自N-末端(氨基末端)中的氨基酸的相应蛋白质合成C-末端(羧基末端)方向。
- 核糖体位于胞质内,或自由漂浮或与内质网有关。
- 它们用来合成蛋白质。
用于蛋白质翻译的核糖体位点
如图所示,每个原核核糖体都有三个trna的结合位点。
- 氨基酰基-TRNA结合位点(或A位点)是在延伸过程中,进入的氨基酰- trna结合的地方。
- 肽基-trna结合位点(或P位点)是与生长多肽链连接的TRNA的结合。
- 退出网站(或E位点)是在翻译中作用的TRNA的结合位点,并且在其从核糖体中释放之前。
这三个位点(A, P和E)都是由核糖体中的rRNA分子形成的。
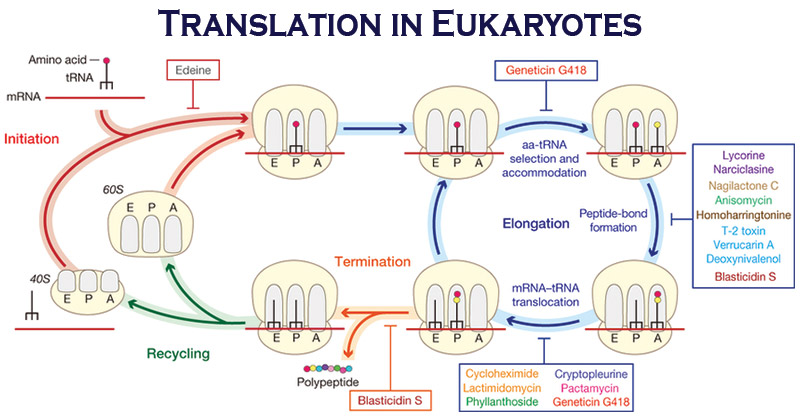
翻译过程
真核生物中蛋白质合成的总机制基本相同原核生物.
然而,有一些显著的差异:
- 原核核糖体的沉降系数为70S,亚基为30S和50S;真核核糖体的沉降系数为80S,亚基为40S和60S。
- 真核核糖体亚基的组成也比原核亚基更复杂,但每个亚单位的功能基本相同,与原核生物中的相同。
- 在真核生物中,每个mRNA是单障碍,即,折扣可能发生的任何后续翻译切割反应;mRNA编码单一蛋白质。在原核生物中,许多MRNA是多排序来编码几种蛋白质。原核mRNA中的每个编码序列都有其自身的起始和终止密码子。
- 与原核生物中的3种起始因子相比,真核生物中蛋白质合成的起始因子至少需要9种不同的真核起始因子。
- 在真核生物中,发起氨基酸是甲硫氨酸,而不是原核生物中的N-甲酰甲基硫氨酸。
- 如在原核生物中,引发需要特殊的引发剂TRNA,并且不同于在mRNA中的内部位置处识别和结合蛋氨酸密码子的TRNA。在准备开始启动的蛋氨酸中加入时,这被称为Met-TRNA我遇见
- 原核生物和真核生物翻译起始的主要区别在于,在细菌中,一个Shine-Dalgarno序列位于AUG起始密码子的5 '处,是30S核糖体亚基的结合位点。
- 相反,大多数真核信使rna不包含Shine-Dalgarno序列。相反,一个40S核糖体亚基附着在mRNA的5 '端并向下移动(即5 '到3 '方向),直到找到AUG起始密码子。这个过程叫做扫描。
- 原核生物的翻译不需要解旋酶,这大概是因为细菌中的蛋白质合成可以在mRNA还在合成的时候就开始了,而在真核生物中,细胞核中的转录和细胞质中的翻译是两个独立的事件,为mRNA二级结构的形成提供了时间。
蛋白质合成(或翻译)进行三个阶段:
- 引发
- 伸长,
- 终止
启动蛋白质合成
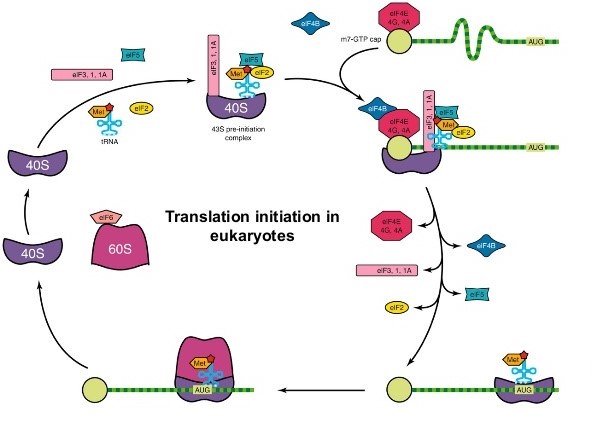
- 第一步是形成一个由40S小核糖体亚基Met-tRNA组成的起始前复合物我遇见、eIF-2和GTP。
- 预发起复合物与真核mRNA的5'末端结合,需要EIF-4F(也称为帽结合复合物)和EIF-3的步骤。
- EIF-4F复合物由EIF-4A,EIF-4E和EIF-4G组成;EIF-4E与MRNA上的5'帽结合,而EIF-4G在聚(a)尾部上的聚(a)结合蛋白相互作用。
- EIF-4A是ATP依赖性RNA螺旋酶,其展开MRNA中的任何二级结构,制备翻译。
- 然后,该复合物在5'至3'方向上沿mRNA移动,直到定位八月启动密码子(即扫描)。
- 真核mrna的5 '非翻译区长度不同,但可以有几百个核苷酸长,可能包含发夹环等二级结构。这些二级结构可能被扫描配合物的起始因子除去。
- 发起密码子通常是可识别的,因为它通常(但并不总是)包含在一个名为的短序列中Kozak共识(5“-ACCAUGG-3”)。
- 一旦复合物被定位在起始密码子上,60S大核糖体亚基结合形成一个80S起始复合物,这一步骤需要GTP的水解并导致几种起始因子的释放。
蛋白质合成伸长
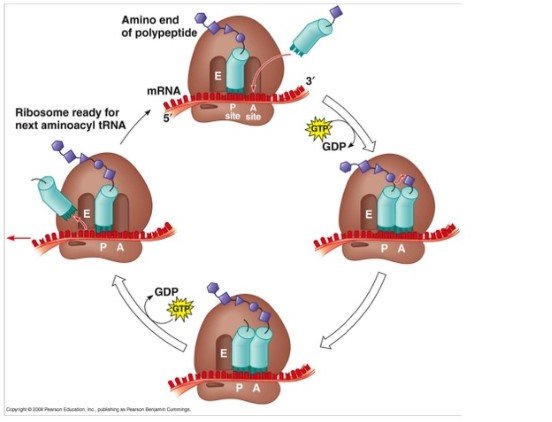
- 延伸取决于真核细胞的延伸因子。
- 涉及三个伸长因子,EEF-1A,EEF-IB和EEF-2,其具有与其原核相提并论EF-TU,EF-TS和EF-G具有相似的功能。
- 在起始步骤结束时,定位mRNA,使得可以在蛋白质合成的伸长阶段翻译下一个密码子。
- 启动子tRNA占据核糖体的P位点,A位点准备接受氨基酰tRNA。
- 在链伸长期间,将每个另外的氨基酸加入到三阶段微循环中的起始多肽链中。
- 这种微循环中的步骤是:
- 将正确的氨基酰基-TRNA定位在核糖体的部位中,
- 形成肽键和
- 通过一个密码子相对于核糖体将mRNA移位。
- 尽管大多数密码子在原核生物和真核生物中编码相同的氨基酸,但在一些真核生物的细胞器内合成的MRNA使用遗传密码的变体。
- 在细菌的伸长过程中,P位点的去乙酰化tRNA在离开核糖体之前移动到E位点。相反,尽管情况还不完全清楚,但在真核生物中,去乙酰化的tRNA似乎是直接从核糖体中喷射出来的。
终止蛋白质合成
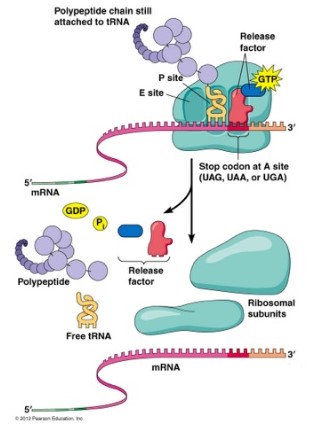
- 伸长率终止取决于真核释放因子。
- 在真核生物中,真核释放因子ERF-1识别所有三个终止密码子(UAA,UAG和UGA),并且在蛋白质ERF-3的帮助下终止翻译。
- 终止时,核糖体被分解,完成的多肽被释放。
参考
- 大卫·海姆斯和奈杰尔·胡珀(2005)。生物化学。第三版。泰勒和弗朗西斯集团:纽约。
- Bailey, W. R., Scott, E. G., Finegold, S. M., & Baron, E. J.(1986)。贝利和斯科特的诊断微生物学。圣路易斯:处于。
- Madigan, M. T., Martinko, J. M., Bender, K. S., Buckley, D. H., & Stahl, D.(2015)。《布罗克微生物生物学》(第十四版)。波士顿:皮尔森。
- https://en.wikipedia.org/wiki/Eukaryotic_translation
你会在真核生物中给我翻译摘要吗?
信息得到了清晰,虽然图表很少,但它们在这个过程中有多么需要